Podjela bakterijskih ćelija. Mehanizam diobe bakterija (prokarioti) Većina bakterijskih stanica dijeli se na svaku
Razmnožavanje bakterija fisijom je najčešći način povećanja veličine mikrobne populacije. Nakon diobe, bakterije narastu do svoje izvorne veličine, za što su potrebne određene tvari (faktori rasta).
Metode razmnožavanja bakterija su različite, ali većina njihovih vrsta ima oblik aseksualne reprodukcije fisijom. Bakterije se retko razmnožavaju pupanjem. Seksualna reprodukcija bakterija prisutna je u primitivnom obliku.
Rice. 1. Fotografija prikazuje bakterijsku ćeliju u fazi diobe.
Genetski aparat bakterija
Genetski aparat bakterija predstavljen je jednim DNK - hromozomom. DNK je zatvoren u krug. Kromosom je lokaliziran u nukleotidu koji nema membranu. Bakterijska ćelija sadrži plazmide.
Nukleoid
Nukleoid je analog jezgra. Nalazi se u centru ćelije. Sadrži DNK, nosilac nasljedne informacije u presavijenom obliku. Nerazmotana DNK doseže dužinu od 1 mm. Nuklearna tvar bakterijske stanice nema membranu, jezgro ili skup kromosoma i ne dijeli se mitozom. Prije podjele, nukleotid se udvostručuje. Tokom diobe, broj nukleotida se povećava na 4.

Rice. 2. Fotografija prikazuje bakterijsku ćeliju u dijelu. U središnjem dijelu je vidljiv nukleotid.
Plazmidi
Plazmidi su autonomni molekuli presavijeni u prsten dvolančane DNK. Njihova masa je znatno manja od mase nukleotida. Unatoč činjenici da su nasljedne informacije kodirane u DNK plazmida, oni nisu vitalni i neophodni za bakterijsku ćeliju.

Rice. 3. Fotografija prikazuje bakterijski plazmid.
Faze podjele
Nakon dostizanja određene veličine karakteristične za odraslu ćeliju, pokreću se mehanizmi diobe.
DNK replikacija
Replikacija DNK prethodi diobi ćelije. Mezozomi (nabori citoplazmatske membrane) drže DNK dok se proces podjele (replikacije) ne završi.
Replikacija DNK se odvija uz pomoć enzima DNK polimeraza. Tokom replikacije, vodonične veze u dvolančanoj DNK se kidaju, što rezultira da se dvije jednolančane kćerke DNK formiraju iz jedne DNK. Nakon toga, kada ćerke DNK zauzmu svoje mjesto u odvojenim ćelijama kćeri, one se obnavljaju.
Čim se replikacija DNK završi, kao rezultat sinteze pojavljuje se suženje, dijeleći ćeliju na pola. Prvo, nukleotid prolazi kroz diobu, zatim citoplazma. Sinteza ćelijskog zida dovršava diobu.
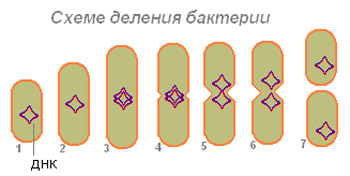
Rice. 4. Šema diobe bakterijskih ćelija.
Razmjena dijelova DNK
Kod Bacillus subtilis, proces replikacije DNK završava se razmjenom dva dijela DNK.
Nakon diobe ćelije formira se most kroz koji DNK jedne ćelije prelazi u drugu. Zatim, oba DNK su isprepletena. Neki dijelovi oba DNK se drže zajedno. Na mjestima adhezije, DNK segmenti se zamjenjuju. Jedna od DNK ide duž skakača natrag u prvu ćeliju.

Rice. 5. Varijanta razmjene DNK kod Bacillus subtilis.
Vrste dioba bakterijskih stanica
Ako je dioba stanica ispred procesa odvajanja, tada se formiraju višećelijski štapići i kokice.
Sinhronom diobom stanica formiraju se dvije pune ćelije kćeri.
Ako se nukleotid dijeli brže od same stanice, tada nastaju multinukleotidne bakterije.
Metode odvajanja bakterija
Podjela razbijanjem
Podjela lomljenjem je karakteristična za bacile antraksa. Kao rezultat ove podjele, stanice se lome na mjestima spajanja, razbijajući citoplazmatske mostove. Zatim se odbijaju, formirajući lance.
Klizna podjela
S kliznim odvajanjem, nakon diobe ćelija se odvaja i, takoreći, klizi duž površine druge ćelije. Ova metoda razdvajanja tipična je za neke oblike Escherichia.
Split split
Sekantom podjelom, jedna od podijeljenih ćelija svojim slobodnim krajem opisuje luk kruga, čiji je centar tačka njegovog kontakta sa drugom ćelijom, formirajući rimski quinque ili klinasti oblik (Corynebacterium diphtheria, Listeria).

Rice. 6. Fotografija pokazuje štapićaste bakterije koje formiraju lance (bacili antraksa).

Rice. 7. Fotografija prikazuje kliznu metodu za odvajanje E. coli.

Rice. 8. Metoda razdvajanja korinebakterija.
Vrsta bakterijskih klastera nakon diobe
Skupine ćelija koje se dijele imaju različite oblike, koji zavise od smjera ravnine podjele.
Globularne bakterije poređani jedan po jedan, dva po dva (diplokoki), u paketima, u lancima ili kao grozdovi. Bakterije u obliku štapa - u lancima.
Bakterije u obliku spirale- haotično.

Rice. 9. Fotografija prikazuje mikrokoke. Okrugle su, glatke, bijele, žute i crvene boje. U prirodi su mikrokoke sveprisutne. Žive u različitim šupljinama ljudskog tijela.

Rice. 10. Na fotografiji su bakterije diplococcus - Streptococcus pneumoniae.

Rice. 11. Fotografija prikazuje bakteriju Sarcina. Kokoidne bakterije skupljaju se u pakete.

Rice. 12. Fotografija prikazuje bakteriju streptokoka (od grčkog “streptos” - lanac). Poređani u lancima. Oni su uzročnici brojnih bolesti.

Rice. 13. Na fotografiji su bakterije "zlatni" stafilokoki. Aranžirani kao "grožđe". Grozdovi su zlatne boje. Oni su uzročnici brojnih bolesti.

Rice. 14. Na fotografiji, namotane bakterije Leptospira su uzročnici mnogih bolesti.

Rice. 15. Fotografija prikazuje štapićaste bakterije iz roda Vibrio.
Stopa podjele bakterija
Stopa podjele bakterija je izuzetno visoka. U prosjeku, jedna bakterijska ćelija se podijeli svakih 20 minuta. U roku od samo jednog dana, jedna ćelija formira 72 generacije potomaka. Mycobacterium tuberculosis se polako dijeli. Cijeli proces podjele traje oko 14 sati.

Rice. 16. Fotografija prikazuje proces diobe ćelija streptokoka.
Seksualno razmnožavanje bakterija
1946. godine naučnici su otkrili seksualnu reprodukciju u primitivnom obliku. U ovom slučaju se gamete (muške i ženske reproduktivne ćelije) ne formiraju, ali neke ćelije razmjenjuju genetski materijal ( genetska rekombinacija).
Kao rezultat toga dolazi do prijenosa gena konjugacija- jednosmjerni prijenos dijela genetske informacije u obliku plazmidi pri kontaktu bakterijskih ćelija.
Plazmidi su mali DNK molekuli. Oni nisu povezani sa genomom hromozoma i sposobni su da se autonomno udvostručuju. Plazmidi sadrže gene koji povećavaju otpornost bakterijskih ćelija na nepovoljne uslove okoline. Bakterije često prenose ove gene jedna na drugu. Primjećuje se i prijenos genetskih informacija na bakterije druge vrste.
U nedostatku pravog seksualnog procesa, konjugacija igra veliku ulogu u razmjeni korisnih karakteristika. Tako se prenosi sposobnost bakterija da ispolje rezistenciju na lijekove. Prijenos otpornosti na antibiotike između populacija koje izazivaju bolesti posebno je opasan za čovječanstvo.

Rice. 17. Fotografija prikazuje trenutak konjugacije dvije E. coli.
Faze razvoja bakterijske populacije
Kada se inokulira na hranjivu podlogu, razvoj bakterijske populacije prolazi kroz nekoliko faza.
Inicijalna faza
Početna faza je period od trenutka sjetve do njihovog rasta. U prosjeku, početna faza traje 1-2 sata.
Faza kašnjenja razmnožavanja
Ovo je faza intenzivnog rasta bakterija. Njegovo trajanje je oko 2 sata. Zavisi od starosti useva, perioda adaptacije, kvaliteta hranljive podloge itd.
Logaritamska faza
Tokom ove faze dolazi do vrhunca u stopi reprodukcije i povećanja bakterijske populacije. Njegovo trajanje je 5 - 6 sati.
Faza negativnog ubrzanja
U ovoj fazi dolazi do pada stope reprodukcije, smanjuje se broj bakterija koje se dijele, a povećava se broj mrtvih bakterija. Razlog negativnog ubrzanja je iscrpljivanje hranjivog medija. Njegovo trajanje je oko 2 sata.
Stacionarna maksimalna faza
U stacionarnoj fazi bilježi se jednak broj mrtvih i novonastalih jedinki. Njegovo trajanje je oko 2 sata.
Faza ubrzanja smrti
Tokom ove faze, broj mrtvih ćelija progresivno raste. Njegovo trajanje je oko 3 sata.
Logaritamska faza smrti
Tokom ove faze, bakterijske ćelije umiru konstantnom brzinom. Njegovo trajanje je oko 5 sati.
Faza smanjenja stope
Tokom ove faze, preostale žive bakterijske ćelije ulaze u stanje mirovanja.

Rice. 18. Slika prikazuje krivulju rasta bakterijske populacije.

Rice. 19. Na fotografiji kolonija Pseudomonas aeruginosa je plavo-zelena, kolonija mikrokoka žuta boja Kolonije Bacterium prodigiosum su krvavo crvene boje, a kolonije Bacteroides niger su crne boje.

Rice. 20. Fotografija prikazuje koloniju bakterija. Svaka kolonija je potomak jedne ćelije. U koloniji, broj ćelija je u milionima. Kolonija raste za 1-3 dana.
Podjela magnetno osjetljivih bakterija
Sedamdesetih godina prošlog stoljeća otkrivene su bakterije koje žive u morima koje su imale osjećaj magnetizma. Magnetizam omogućava ovim nevjerovatnim stvorenjima da se kreću duž linija magnetsko polje Zemlju i pronađite sumpor, kiseonik i druge supstance koje su joj toliko potrebne. Njihov "kompas" predstavljaju magnetosomi, koji se sastoje od magneta. Kada se dijele, magnetski osjetljive bakterije dijele svoj kompas. U tom slučaju, stezanje tijekom diobe postaje očito nedovoljno, pa se bakterijska ćelija savija i pravi oštar prijelom.

Rice. 21. Fotografija prikazuje trenutak podjele magnetno osjetljive bakterije.
Rast bakterija
Kada bakterijska ćelija počne da se deli, dva molekula DNK kreću se na suprotne krajeve ćelije. Zatim se ćelija dijeli na dva jednaka dijela, koji su odvojeni jedan od drugog i povećavaju se na svoju prvobitnu veličinu. Brzina podjele mnogih bakterija je u prosjeku 20 - 30 minuta. U roku od samo jednog dana, jedna ćelija formira 72 generacije potomaka.
Tokom procesa rasta i razvoja, masa ćelija brzo apsorbuje hranljive materije iz okoline. Tome doprinose povoljni faktori okoline - temperaturni uslovi, dovoljne količine hranljivih materija i potreban pH sredine. Aerobne ćelije zahtevaju kiseonik. Opasno je za anaerobe. Međutim, u prirodi se ne događa neograničeno razmnožavanje bakterija. Sunčeva svjetlost, suv zrak, nedostatak hrane, visoka temperatura okoline i drugi faktori štetno djeluju na bakterijsku ćeliju.

Rice. 22. Fotografija prikazuje trenutak diobe ćelije.
Faktori rasta
Za rast bakterija neophodne su određene supstance (faktori rasta), od kojih neke sintetiše sama ćelija, a neke dolaze iz okoline. Potreba za faktorima rasta je različita za sve bakterije.
Potreba za faktorima rasta je stalna karakteristika, što omogućava da se koristi za identifikaciju bakterija, pripremu hranljivih podloga i upotrebu u biotehnologiji.
Faktori rasta bakterija (bakterijski vitamini) su hemijski elementi, od kojih su većina vodotopivi vitamini B. U ovu grupu spadaju i hemin, holin, purinske i pirimidinske baze i druge aminokiseline. U nedostatku faktora rasta dolazi do bakteriostaze.
Bakterije koriste faktore rasta u minimalnim količinama i nepromijenjene. Brojne hemikalije u ovoj grupi su deo ćelijskih enzima.

Rice. 23. Fotografija prikazuje trenutak podjele bakterije u obliku štapa.
Najvažniji faktori rasta bakterija
- vitamin B1 (tiamin). Učestvuje u metabolizmu ugljenih hidrata.
- Vitamin B2" (riboflavin). Učestvuje u redoks reakcijama.
- Pantotenska kiselina je komponenta koenzima A.
- vitamin B6 (piridoksin). Učestvuje u metabolizmu aminokiselina.
- Vitamini B12(kobalamini su supstance koje sadrže kobalt). Oni aktivno učestvuju u sintezi nukleotida.
- Folna kiselina. Neki od njegovih derivata su dio enzima koji katalizuju sintezu purinskih i pirimidinskih baza, kao i nekih aminokiselina.
- Biotin. Učestvuje u metabolizmu dušika i također katalizuje sintezu nezasićenih masnih kiselina.
- Vitamin PP(nikotinska kiselina). Učestvuje u redoks reakcijama, stvaranju enzima i metabolizmu lipida i ugljikohidrata.
- Vitamin H(para-aminobenzojeva kiselina). To je faktor rasta mnogih bakterija, uključujući i one koje nastanjuju ljudska crijeva. Folna kiselina se sintetizira iz para-aminobenzojeve kiseline.
- Gemin. Sastavni je dio nekih enzima koji sudjeluju u reakcijama oksidacije.
- Kholin. Učestvuje u reakcijama sinteze lipida ćelijskog zida. Dobavljač je metil grupe u sintezi aminokiselina.
- Purinske i pirimidinske baze(adenin, gvanin, ksantin, hipoksantin, citozin, timin i uracil). Supstance su potrebne uglavnom kao komponente nukleinskih kiselina.
- Amino kiseline. Ove supstance su komponente ćelijskih proteina.
Potreba za faktorima rasta određenih bakterija
Auksotrofi Da bi osigurali život, zahtijevaju snabdijevanje kemikalijama izvana. Na primjer, klostridije nisu u stanju sintetizirati lecitin i tirozin. Stafilokoki zahtijevaju opskrbu lecitin i arginin. Streptokoki zahtijevaju opskrbu masnim kiselinama - komponentama fosfolipida. Corynebacteria i Shigella zahtijevaju nikotinsku kiselinu. Staphylococcus aureus, pneumokoke i Brucella zahtijevaju vitamin B1. Bacili streptokoka i tetanusa - u pantotenskoj kiselini.
Prototrofi samostalno sintetizirati potrebne tvari.

Rice. 24. Različiti uslovi okoline imaju različite efekte na rast kolonija bakterija. Na lijevoj strani je stalan rast u obliku kruga koji se polako širi. Desno je brzi rast u obliku "izbojaka".
Proučavanje potrebe bakterija za faktorima rasta omogućava naučnicima da dobiju veliku mikrobnu masu, toliko potrebnu u proizvodnji antimikrobnih lijekova, seruma i vakcina.
Više o bakterijama pročitajte u člancima:
Proliferacija bakterija je mehanizam za povećanje broja mikrobnih populacija. Bakterijska dioba je glavni način razmnožavanja. Nakon dijeljenja, bakterije moraju dostići veličinu odrasle osobe. Bakterije rastu tako što brzo apsorbuju hranljive materije iz svog okruženja. Za rast su potrebne određene tvari (faktori rasta), od kojih neke sintetizira sama bakterijska stanica, a neke dolaze iz okoline.
Proučavajući rast i razmnožavanje bakterija, naučnici neprestano otkrivaju korisne karakteristike mikroorganizmi, čija upotreba u Svakodnevni život a u proizvodnji je ograničena samo njihovim svojstvima.
U završnoj fazi diobe, ćelijska membrana se skuplja i razgrađuje ili učestvuje u sintezi septuma nakon čega slijedi autoliza, formirajući dvije odvojene ćelije
Proces diobe u bakterijama kontrolira homolog tubulina, protein FtsZ, koji formira prstenastu strukturu na mjestu diobe.
Zajedno sa FtsZ, na mjestu diobe formira se skup od otprilike 8 proteina uključenih u diobu
Mjesto diobe ćelije određuju dva negativna regulatorna sistema: blokirajući efekat nukleoida i Min sistem
Većina ćelija eukarioti podijeliti tačno po sredini, formirajući dvije identične ćelije kćeri. Podjela je usklađena sa završetkom replikacije i segregacije hromozoma. Tipično, do diobe dolazi na kraju perioda rasta, tokom kojeg se ćelijska masa udvostručuje. Nakon segregacije hromozoma dolazi do citokineze, uslijed čega se stanica dijeli na dva dijela. Tokom citokineze, svi slojevi ćelijske membrane lokalno poprimaju prstenasti oblik. Kao što je prikazano na donjoj slici, citokineza se javlja na najmanje dva različita načina.
U gram-negativnim mikroorganizmi, kao što je E. coli, do diobe dolazi kontrakcijom slojeva postojeće membrane, nakon čega slijedi pucanje nastalog mosta. Kod drugih bakterija, kao što je gram-pozitivna B. subtilis, novoformirane prstenaste strukture materijala stanične stijenke rastu u ćeliju, formirajući septum. Kada je formiranje septuma završeno, između sestrinskih ćelija formira se dvostruka membrana, ali ćelije ostaju povezane jedna s drugom. Podjela ćelije je nezavisan događaj koji uključuje autolizu septalnog materijala. U zavisnosti od uslova rasta, autoliza septuma se može odvijati prilično sporo i biti praćena pojavom dugih lanaca međusobno povezanih ćelija.
Prilikom isticanja i karakterizacije fis mutants(filamentne temperaturno osjetljive mutacije), identificiran je niz gena potrebnih za fisiju. mutantne ćelije fis na temperaturama koje nisu dopuštene rastu u obliku dugih filamenata koji se ne dijele. Oko 8 fis gena nalazi se u većini bakterija. Lutkenhaus je napravio plodno zapažanje, koji je otkrio da protein FtsZ formira prstenaste strukture neposredno ispod ćelijske membrane na mjestu diobe. Zatim se preostali proteini diobe približavaju ovom "Z-prstenu" određenim redoslijedom. Ovaj proces za ćelije E. coli prikazan je na slici ispod. Funkcije većine ovih proteina su nepoznate.
Ključne podjele proteina FtsZ, je homolog eukariotskog tubulina, proteina koji je dio citoskeleta i formira mikrotubule. Poput tubulina, ovaj protein je GTPaza i, u prisustvu GTP-a, polimerizira se da formira linearne proto-filamente koji formiraju snopove i planarne strukture in vitro. Prstenasta struktura proteina FtsZ je izuzetno dinamična i in vivo je stalno podložna reformaciji (sa poluživotom<10 с!). В этом отношении белок напоминает тубулин эукариот.
Kod prokariota, podjela se događa ili stvaranjem suženja ili septuma.Radi jednostavnosti, kapsularni sloj ćelijske membrane nije prikazan.
U Z-prstenu sa proteinom FtsZ FtsA protein je u direktnoj interakciji, čija je funkcija vjerovatno da stabilizira prsten. FtsA protein podsjeća na aktin u eukariotskim stanicama, ali ima dodatni domen čija je funkcija nepoznata. Ovaj protein stvara dimere, ali se ne čini da polimerizira. Iako nije uključen u formiranje Z-prstena, dvostruke mutantne stanice s manjkom FtsA i ZipA proteina nisu u stanju da formiraju prstenaste strukture. Dakle, funkcije FtsA i ZipA proteina se djelomično preklapaju, a barem jedna od njih je potrebna za stabilizaciju Z-prstena. Takođe je pokazano da ZipA protein direktno interaguje sa FtsZ i, za razliku od potonjeg i FtsA, je transmembranski protein. Stoga ZipA može posredovati u konjugaciji Z-prstena sa ćelijskom membranom.
Odmori se fisioni proteini su transmembranski proteini. Funkcije FtsL i FtsQ proteina su nepoznate. FtsW protein vjerovatno opskrbljuje prekursore za protein FtsI, koji je enzim uključen u sintezu septuma. Potonji ima sposobnost da veže penicilin i stupa u interakciju sa aparatom za sintezu ćelijskog zida koji funkcioniše tokom deobe. Proteini FtsK i FtsN neophodni su za diobu ćelija u E. coli, međutim, kod B. subtilis, homolog proteina FtsK (SpoIIIE) nije uključen u diobu i nema homologa FtsN proteina u ovim stanicama.
Između dva dobro proučeno mikroorganizmi, E. coli i B. subtilis, postoje zanimljive razlike u sastavljanju fisionih proteina. Tako je kod E. coli ovaj proces gotovo linearan, dok je kod B. subtilis sklapanje proteina na strukturi Z-prstena međuzavisno. Ove razlike vjerovatno odražavaju različitu organizaciju ćelijskog zida između Gram-negativnih i Gram-pozitivnih mikroorganizama. Još uvijek znamo malo o tome kako potpuno sastavljeni aparat za podjelu utječe na citokinezu, a razjašnjavanje ovih pitanja predstavlja ogromno polje aktivnosti za istraživače.
Divizija se kontroliše uglavnom na nivou formiranje FtsZ prstena. Pretpostavlja se da položaj mesta podele, a verovatno i tok ovog procesa tokom vremena kontrolišu dva faktora: blokiranje nukleoidom i Min sistem. Oba ova faktora osiguravaju početak diobe tek nakon što se završi replikacija DNK, kao i ista veličina rezultirajućih ćelija.
Blokirajući faktor nukleoid nedovoljno istražen. Ona se manifestuje u tome što nukleoid zbog svog volumena može spriječiti diobu. Stoga se dioba ćelije događa tek nakon završetka kruga replikacije DNK i divergencije sestrinskih hromozoma kako bi se formirali pojedinačni nukleoidi. Kada su procesi replikacije ili segregacije blokirani, prisustvo nukleoida u sredini ćelije sprečava stvaranje septuma. U principu, negativan efekat nukleoida može se jednostavno objasniti odsustvom FtsZ proteina u ovoj regiji. U ovom slučaju, protein se ne akumulira do kritične koncentracije potrebne za njegovu polimerizaciju.
Značaj faktor blokiranja nukleoida Potencijalni problem za ćeliju je taj što stanični polovi (barem kod bakterija u obliku štapa) nisu zaštićeni nukleoidom, pa stoga može doći do aberantne polarne podjele. Da bi se to spriječilo, mnoge bakterije sadrže proteine koji su dio Min sistema, koji sprječava diobu na polovima.Naziv ovog sistema potiče od naziva mini-ćelija koje formiraju mini-mutanti, a koje karakterizira dioba na polovima. stubovi.
Ključ sistem efektor Min je inhibitor ćelijske diobe nazvan MinC. Ovaj protein ima sposobnost da inhibira formiranje Z-prstena, vjerovatno direktnim inhibiranjem FtsZ polimerizacije. MinC aktivnost kontroliše MinD protein. Ovaj protein vjerovatno kontrolira subcelularnu lokalizaciju MinC kroz dva različita mehanizma. Jedna od njih je da MinD prenosi MinC do periferije ćelije (bliže citoplazmatskoj membrani) gdje se sklapa FtsZ prstenasta struktura. Drugi mehanizam je da MinD ograničava aktivnost MinC na ćelijske polove, čime se sprječava početak polarne diobe, ali promoviše dioba stanica na srednjoj liniji.
Mnoge bakterije u obliku štapa imaju sistem MiniCD koristi se za kontrolu lokacije mjesta podjele. Ovaj sistem je dobro okarakterisan kod bakterija E. coli i B. subtilis. Zanimljivo je da ova dva mikroorganizma imaju veoma različite mehanizme pomoću kojih MinD ograničava efekat MinC na polovima ćelije. B. subtilis koristi jednostavan mehanizam u kojem polarni sidreni protein DivIVA transportuje MinCD kompleks do polova ćelije i tamo ga drži u statičkom položaju tokom čitavog ćelijskog ciklusa. Kao što je prikazano na donjoj slici, DivIVA i MinD se lokalizuju na polovima novoformirane ćelije, a prisustvo inhibitora MiniC sprečava formiranje FtsZ prstena na polovima.
Očigledno nakon što je replikacija završena DNK, novo potencijalno mjesto diobe se kreira u sredini ćelije. Koncentracija MiniC inhibitora na polovima omogućava sklapanje FtsZ prstena u sredini ćelije i osigurava mobilizaciju drugih proteina diobe. U ovom trenutku, aparat za podjelu vjerovatno postaje neosjetljiv na inhibitorni efekat MinC.Tada se proteini DivIVA i MinD kreću u sredinu ćelije. Stoga, kada se tokom diobe formira novi par ćelijskih polova, DivIVA se ubacuje u nove polove i formira novo mjesto za ekspresiju inhibitornog efekta MinCD. Kada dođe do kontrakcije ljuske, dolazi do rasklapanja FtsZ prstena, ali DivIVA i MinCD ostaju na novoformiranim polovima, čime se sprečava podelu na ovim polarnim mestima.
Dakle, prevoz DivIVA do mjesta diobe i njegovo naknadno zadržavanje na polovima ćelije ključni su događaji u ovom mehanizmu.
Zanimljivo, proteini DivIVA lokalizira se na mjesta diobe kada se eksprimira u eukariotskim stanicama (fisijski kvasac). Ovo sugerira da DivIVA može prepoznati topološke karakteristike, kao što je zakrivljenost membrane, a ne specifične proteinske mete.
Za razliku od ovoga, u ćelijama E. coli postoji dinamički sistem MinCD, koji neko vreme skuplja kompleks na jednom polu. Zatim se rastavlja i ponovo sastavlja na suprotnom polu. Ovo se ponavlja mnogo puta. Ovaj proces kontroliše MinE proteinski prsten, koji se, zauzvrat, svaki put pomiče na jedan ili drugi pol, istiskujući MinCD i omogućavajući mu da se skupi na suprotnom polu. Promjene u lokalizaciji MinCD-a od jednog do drugog pola se dešavaju s frekvencijom od nekoliko desetina sekundi. Kao što je prikazano na donjoj slici, MinD se naizmjenično akumulira na periferiji membrane sa svake strane MinE prstena. Brza relokalizacija MinD-a sprečava da se FtsZ prsten sklopi na polovima.
Prisutnost MinE u centralnom regionu eliminiše manifestaciju inhibitornog efekta MinD-a i dozvoljava FtsZ prstenu da se sklopi na ovom mestu. Ostaje nejasno zašto je E. coli razvila tako energetski nepovoljan mehanizam za kontrolu MinCD i uspostavljanje polova.
MinD pripada zanimljivoj grupi proteina koji dijele zajedničku funkciju vezivanja nukleotida, što također uključuje protein za razdvajanje hromozoma, ParA. Protein blizak ParA, Soj, takođe pokazuje dinamička svojstva. Ono što ovi proteini vjerovatno imaju zajedničko je njihova sposobnost da vežu i hidroliziraju nukleotide i kontroliraju reakcije polimerizacije i depolimerizacije. Ovo podsjeća na mehanizam koji kontrolira dinamičku nestabilnost aktinskih filamenata i mikrotubula kod eukariota. Stoga ovi proteini pripadaju drugoj klasi bakterijskih proteina citoskeleta koji imaju široke funkcije koje se posebno odnose na pitanja morfogeneze u različitim fazama ćelijskog ciklusa.
Nedavno gram-pozitivne bakterije identificiran je protein uključen u blokiranje diobe stanica od strane nukleoida. Ovo je Noc, koji je protein koji se nespecifično veže za DNK i lokaliziran je u nukleoidu. Takođe je inhibitor deobe ćelija. Ako replikacija hromozoma nije poremećena, noc mutanti rastu normalno. Štaviše, u noc-ćelijama se dioba događa uz učešće nukleoida, dok se ćelije divljeg tipa ne dijele. Kao što je prikazano na slici ispod, Noc i MiniCD sistem lociraju FtsZ prsten u sredini ćelije. U ćelijama divljeg tipa, DivIVA pokreće polimerizaciju MinD proteina, koji se širi od polova do sredine ćelije duž membrane.
Protein MinC vezani za proteine MinD, sprečava FtsZ od akumulacije ili polimerizacije u blizini polova ćelije. Pretpostavlja se da se Noc protein veže za nukleoid i inhibira akumulaciju FtsZ ili ispoljavanje njegove aktivnosti u blizini nukleoida. U noc-ćelijama, Min sistem sprečava sklapanje FtsZ prstena, isključujući region srednje ćelije, i ćelije rastu normalno. Međutim, u min ćelijama, Noc inhibira sklapanje FtsZ samo oko nukleoida, a FtsZ formira prstenastu strukturu u sredini ćelije i na polovima gde nema nukleoida. U ćelijama kojima nedostaju topološki inhibitori (min-noc-dvostruki mutanti), sklapanje FtsZ je neometano i brojne zakrpe ovog proteina se formiraju u celoj ćeliji. Njihovo formiranje dovodi do gubitka sposobnosti ćelije da se deli. Gram negativnim bakterijama nedostaje Noc, ali E. coli ima protein koji kontroliše sistem za blokiranje podjele nukleoida mehanizmom sličnim Noc.
Bakterijski organizmi su dugo kolonizirali sva poznata staništa. Oni su u vazduhu, u vodi i žive u drugim organizmima. Ali većina ih je u gornjim slojevima tla. Broj ovih organizama ne zavisi samo od strukturnih karakteristika. Višestruko se povećava zbog svoje velike sposobnosti reprodukcije. Kako se bakterije razmnožavaju bit će ukratko opisano u članku.
Šta su bakterije?
Ovi organizmi su jednoćelijski, rjeđe kolonijalni organizmi. Oni su raspoređeni prilično primitivno. Površinski aparat predstavljen je membranom i mukoznom kapsulom, a citoplazma je lišena mitohondrija i plastida. Mnoge ćelije imaju flagelum pomoću kojih se bakterija može kretati.
Genetski materijal
Bakterije su prokarioti. To znači da njihovim ćelijama nedostaje jezgro. Ali genetski materijal je još uvijek prisutan u njima. Klasteri molekula DNK nalaze se u određenom dijelu citoplazme i nazivaju se nukleoidi. Drugim riječima, možemo reći da prokarioti imaju jezgro bez ljuske. Zbog toga ne mogu obavljati složene biohemijske procese. Međutim, to ni na koji način ne utječe na njihovu sposobnost reprodukcije.
Kako se razmnožavaju bakterije?
Bakterije se razmnožavaju Ovo je glavni i najbrži način. Od jedne matične ćelije za pola sata nastaju dve ćerke ćelije. I nakon istog vremenskog perioda, nove ćelije se ponovo formiraju od dve ćelije kćeri. Ovo objašnjava veliki broj bakterija u prirodi.
U nepovoljnim uslovima, bakterije su u stanju da formiraju spore - ćelije povremeno pupolje - formiraju male izbočine koje rastu, pretvaraju se u odrasle jedinke i odvajaju se od majke.

Kako se bakterije razmnožavaju može se također razmotriti na primjeru konjugacije. Ovo je oblik seksualnog procesa. Sastoji se u razmjeni nasljednih informacija između ćelija. Prije početka, kružni molekul DNK se udvostručuje. Zatim se između stanica formira citoplazmatski most, duž kojeg se jedna stanica kreće u drugu. Postoji razmjena DNK dijelova. Kao rezultat, tijelo dobiva nove karakteristike koje su mu najčešće korisne. Na primjer, bakterije postaju otporne na štetne faktore okoline, viruse ili antibiotike.
Žive i razmnožavaju se na korijenu mahunarki i žitarica. Prodirući u korijenski sistem kroz zahvaćena područja ili korijenske dlake, rastu i formiraju izbočine - nodule. U njima se stvara povoljno okruženje za metabolizam. Korijen daje bakterijama organske tvari, a bakterije dušik koji je toliko neophodan za rast i razvoj biljaka.
Podela ćelije na dva dela
Način na koji se bakterije razmnožavaju ovisi o njihovoj vrsti i staništu. Ali svi bakterijski organizmi su sposobni da se podijele na dva dijela. Ovaj proces se odvija u nekoliko faza i naziva se binarna fisija.
Prije nego što podjela počne, kružni molekul DNK se udvostručuje. Drugim riječima, dolazi do replikacije. Nukleotid se dijeli, a ćerke DNK se razilaze. Rastući u citoplazmu, ćelijska membrana se nalazi između molekula DNK. Ona je ta koja dijeli ćeliju i njen sadržaj na pola.
72 bakterijske generacije se rađaju iz jedne ćelije dnevno. Kada bi sve ove bakterije ostale održive, njihova biomasa bi bila oko 5 tona.To se naravno ne dešava u prirodi i većina bakterija umire.
Vegetativno razmnožavanje
Struktura također određuje kako se bakterije razmnožavaju.
Kolonijalne vrste i cijanobakterije (plavo-zelene alge) su sposobne za vegetativnu reprodukciju. Biljke se najčešće razmnožavaju na ovaj način. Sastoji se od odvajanja njegovog višećelijskog dijela od cijelog organizma.

Filamentne vrste cijanobakterija formiraju specijalizirane ćelije koje se nazivaju heterociste. Vegetativno razmnožavanje sastoji se od lomljenja filamenata čija granica prolazi na mjestu heterocista.
Koke mogu formirati lance, grozdove ili druge formacije. Odvajajući se jedno od drugog, oni se takođe množe.
Sporulacija
Bakterije se razmnožavaju sporama koje nastaju kada se pojave nepovoljni uslovi. Sporulacija nije samo metoda razmnožavanja. Unutar spore se stvara posebno okruženje, sadržaj vode se smanjuje, a vitalni procesi se zaustavljaju. U ovom stanju, spore se ne boje visokih temperatura, jonizujućeg zračenja ili izlaganja hemikalijama. Kada se vrate povoljni uslovi, iz spora izlaze mladi bakterijski organizmi. Stoga je stvaranje spora dodatna prilika da se održi vitalnost ćelija u uslovima neprikladnim za život. Postoje slučajevi kada su bakterijske spore ostale održive desetinama, pa čak i stotinama godina.

encistiranje
Drugi način zaštite od nepovoljnih uslova i način razmnožavanja je stvaranje cista. To su mehurići sa debelim školjkama. Bakterije mogu dugo ostati u stanju ciste. Međutim, ne umiru od temperatura koje prelaze 200 stepeni. Sa početkom normalnih uslova, bakterija napušta ljusku i počinje normalnu binarnu fisiju.

Način na koji se bakterije razmnožavaju ovisi o uvjetima okoline. Kada nema dovoljno hranljivih materija i vlage, postoji višak kiseonika, vazduh je previše vruć ili hladan, bakterije koriste procese encistiranja ili sporulacije. U ugodnim uvjetima dijele se ili razmnožavaju vegetativno. Upravo ta raznolikost metoda reprodukcije za koje su bakterije sposobne određuje njihov broj u prirodi. Ako se proces podjele jedne bakterijske ćelije ne zaustavi 10 dana, mogle bi pokriti cijelu površinu zemaljske kugle.
Bakterije su najstariji oblik života na Zemlji. Pojavili su se na planeti prije otprilike 3,8-3,6 miliona godina. Agresivni klimatski uslovi učinili su ih izdržljivim i otpornim na preživljavanje. Najstarija stvorenja bi bile cijanobakterije.
Oni su doprinijeli akumulaciji kisika u atmosferi. Naše tijelo se sastoji od brojnih vrsta njih. Postoje korisne i štetne vrste. Žive svuda: u vodi, u vazduhu, u ljudima i životinjama, u slojevima tla.
Volumen kolonija ovisi ne samo o strukturi, već i o tome kako se bakterije dijele. Struktura je primitivna. Čini se da je uređaj mukozna kapsula ili membrana. Mikroorganizam se sastoji od samo jedne žive ćelije.
U citoplazmi nema mitohondrija ili plastida. Većina mikroba ima flagele i antene, uz pomoć kojih se kreću kroz krv, krvne sudove i tkiva. Oni su prokarioti, što znači da nemaju jezgro.
To znači da se mikročestice DNK akumuliraju u određenom dijelu citoplazme. Zovu se nukleotidi. Nukleotidi su neka vrsta jezgra; oni sadrže informacije. DNK pohranjuje informacije u komprimiranom obliku. Kada se rasklopi, dužina doseže 1 mm.
Bakterije se razmnožavaju fisijom.
Trebali biste znati da se bakterije razmnožavaju samo uz prisustvo povoljnih faktora, koje ćemo razmotriti u nastavku.
Za njihov rast potrebno je:
- svjetlo;
- temperatura;
- prisustvo kiseonika;
- vlažnost;
- faktor alkalnosti i kiselosti;
Temperaturni uslovi zanimaju ljekare. Da bi se ćelije podijelile potrebna je određena temperatura. Neke klase, na vrlo niskim nivoima, padaju u stanje suspendirane animacije ili hibernacije, dok druge, samo na visokim nivoima, ne mogu nastaviti svoj rast i bivaju uništene.
Dok se neki mogu ubiti ključanjem vode, drugi se sasvim dobro snalaze sa smrzavanjem. Među ovim ograničenjem postoje prosječni uvjeti pod kojima se maksimalni razvoj može dogoditi velikom brzinom. Potrebna temperaturna faza je od 23 do 30 stepeni, za protok patogene flore potrebno je 38 stepeni.
Bakterijske protozoe napreduju u ovom okruženju. U idealnim uslovima, prokarioti su u stanju da proizvedu 34 triliona potomaka dnevno. Stanje odrastanja nastupa za oko 20 minuta. Srećom, ne žive dugo, nekoliko minuta ili sati.
Šta je potrebno nekim mikroorganizmima?

Grupi stafilokoka potrebni su arginin i lecitin. Streptokoki u fosfolipidima. Shigella i corina bakterije trebaju nikotinsku kiselinu da bi se hranile. Staphylococcus aureus, pneumokok i bruceloza ne mogu živjeti bez vitamina B1, ali prototrofi sami sintetiziraju ono što im je potrebno.
Putevi sazrevanja

Kao što je ranije spomenuto, razvoj protozoa se odvija kroz diobu.
Dešava se:
- jednostavno;
- pupanje;
- konjugacija, seksualni odnos;
Najlakši način
S prvom metodom, bakterije se mogu razmnožavati jednakom poprečnom podjelom. Matične ćelije, nakon udvostručavanja lanaca DNK i organela, formiraju dva dijela, odnosno ćelije kćeri. Genetski kod se formira slično kao i kod majke.
Kao da se kloniraju. U roku od 24 sata iz jedne ćelije izlazi 70 generacija. Ako pretpostavimo da su svi mogli živjeti, masa je bila veća od 5 tona. Naravno, to je u prirodi nemoguće.
Vegetativna faza
Ili, jednostavnije, na pupanje ukazuje činjenica da stvorenja izrastu drugi pupoljak na jednom od polova, to jest, sami sebe. Kada dođe do grananja, lanci DNK pucaju. U procesu učestvuju heterociste. Ovu metodu koriste cijanobakterije i kolonijalne stijene.
Na taj način prokarioti mogu narasti do 4 pupoljka, nakon čega dolazi do starenja i smrti. Kolonije koka se odvajaju i slobodno rastu.
Sporulacija

Postoji bifurkacija sporova.
Kako se to dešava?
Bacili se na taj način razmnožavaju kada nastupe nepovoljni vanjski i unutrašnji uslovi okoline. Unutar spore se stvara posebno okruženje, mehanizam života je suspendovan, a nivo vode opada. Ako bacil dođe u takvo stanje, ne plaši se ni hladnoće, ni vrućine, ni zračenja različite etiologije, ni hemikalija.
Čim se faktori poboljšaju, pojavljuju se mladi prokarioti. Ciklus postaje veoma dug. Nauka čak zna za slučajeve kada su naučnici pronašli protozoe stare desetine ili čak stotine godina.
Seksualni trakt

Konjugacija se javlja kod bakterija koje žive prvenstveno u ljudskom ili životinjskom tijelu. Ovdje dva oblika dolaze u kontakt jedan s drugim i počinje razmjena podataka. To se zove genetska rekombinacija, formiranje novih vrsta.
E. coli bakterije i drugi gram-pozitivni i gram-negativni tipovi se razmnožavaju spolno. Ako nema pravog smjera, onda je takva razmjena među njima korisna i može doprinijeti razvoju rezistencije na antibiotike i druge lijekove.
encystation

Drugi način zaštite od agresivnih okolnosti je transformacija u ciste. Ciste se odnose na vezikule u debeloj membrani. Bacili mogu ostati u ovom položaju veoma dugo. Ni 200 stepeni Celzijusa ih neće uništiti. Dalje, iz pozitivnih razloga, oni izlaze dijeljenjem binarno.
Dakle, metode umnožavanja patogena podliježu vanjskom okruženju. Nedostatak vode, visok sadržaj kiseonika u vazduhu, nedostatak visoko hranljivih mikroelemenata. Promjene niske ili visoke temperature prisiljavaju čovjeka da pribjegne sporulaciji, encistaciji.
Obim bakterijske populacije

Živeći u povoljnim uslovima, ćelije su u početnoj fazi, početnoj fazi. Prosječno trajanje 1-2 sata. Inhibicija rasta traje oko nekoliko sati. Sa logaritamskim periodom, bacili se mogu brzo razmnožavati, dostižući vrhunac nakon 6 sati.
Negativno ubrzanje kada su nutritivne rezerve mikroelemenata i supstanci iscrpljene. Stacionarna faza, mrtve jedinke se zamjenjuju novim nakon dva sata. Faza ubrzane smrti, bacili umiru svaka 3 sata. Logaritamska faza, obilježena trajnom smrću, traje 6 sati.
Stopa smrti se smanjuje; u ovom trenutku preostale žive ćelije ulaze u stanje mirovanja.
Višećelijska faza

Jednoćelijska faza je sposobna obavljati sve funkcije tijela, na to ne utječu susjedni mikroorganizmi. Jednoćelijski organizmi formiraju ćelijske agregate, drže ih zajedno sluz.
Često se nakupina bacila pojavljuje u jednoj grani. Na taj način mikobakterije razvijaju ciste, što rezultira svojevrsnom razmjenom. Ovaj fenomen služi kao preteča višećelijske formacije. To uključuje cijanobakterije i aktinomicete.
Koje uslove pojedinci moraju ispuniti:
- agregacija ćelija;
- podjela imovine između njih;
- uspostavljanje pravilnog kontakta između pojedinaca;
Kod filamentoznih pojedinaca, struktura je opisana u ćelijskom zidu, stvarajući odnose između pojedinaca. Bakterije razmjenjuju tvari i energiju. Neke filamentne vrste, pored vegetativnih jedinki, sadrže diferencijalne heterociste ili akinete.
Lokalizacija
Ovisno o razgradnji, bacili imaju određene vrste nakupina:
- sferni;
- spirala;
Prvi se nalaze u paru ili jedan po jedan; to su diplokoki, mikrokoki i stafilokoki. Mogu izgledati kao grane grožđa ili lanci. U obliku spirale, rasuti na haotičan način, to uključuje leptospirozu i vibrio.
Neki mikroorganizmi se razmnožavaju sporulacijom (aktinomicete i gljive) i pupanjem (kvasac); neki mikroorganizmi se razmnožavaju spolno, ali većina ih se razmnožava aseksualno (vegetativno). Pod povoljnim uslovima, reprodukcija se odvija izuzetnom brzinom - svakih 20-30 minuta matična bakterijska ćelija se deli na dve ćelije kćeri. Ćerka ćelija na kraju postaje matična ćelija i takođe se deli. Dakle, podjela bakterija se odvija geometrijskom progresijom. Ako bi se takva podjela odvijala nesmetano, onda bi nakon 48 sati jedna bakterija mogla dati nastanak stotina milijardi ćelija, a nakon pet dana takvu masu koja bi ispunila bazene svih mora i okeana. Međutim, to se ne događa, jer na mikroorganizme utječu različiti faktori okoline.
Ćelijskoj diobi prethodi ravnomjerno povećanje ukupnog dušika, RNK i proteina u citoplazmi. Tada dolazi do replikacije DNK (udvostručavanja). U ćeliji koja se dijeli, vodonične veze se prekidaju između spirala DNK i formiraju se jednostruke ćerke DNK spirale (slika 25).
Rice. 25. Proces binarne fisije štapićastih prokariota
3 - istezanje ćelije;
- formiranje septuma;
5 - razdvajanje ćelija.
Neposredno nakon replikacije DNK, počinje produžavanje ćelije i formiranje poprečnog septuma zbog dva sloja citoplazmatske membrane koji strše jedan prema drugom. Najčešće se u sredini matične ćelije formira septum, zbog čega ćelije kćeri imaju približno istu veličinu. Između slojeva septuma formira se ćelijski zid.
Tokom procesa reprodukcije, jedna od polovica ćelije stalno zadržava flagele. U završnoj fazi razmnožavanja bakterija, flagele rastu u drugoj polovici.
Rast i razmnožavanje mikroorganizama zavisi od različitih faktora životne sredine i karakteristika vrste. Posmatranje razvoja mikroorganizama uzgajanih u tečnom hranljivom mediju u zatvorenim rezervoarima pokazuje da je za rast biomase potrebno prisustvo izvora energije, prisustvo komponenti neophodnih za sintezu biomase, odsustvo inhibitora u medijumu koji potiskuju rast ćelija i održavanje u okruženju potrebnih fizičkih i hemijskih uslova. Pod ovim uslovima, rast mikroorganizama se može podeliti u nekoliko uzastopnih faza ili perioda (slika 26):

1. lag faza (engleski lag - kašnjenje) - period između sjetve bakterija i početka razmnožavanja. Tokom ovog perioda, bakterijska kultura se prilagođava hranljivom mediju. Manifestuje se u akumulaciji optimalne količine potrebnih enzima, u inaktivaciji nekog inhibitora prisutnog u medijumu, u klijanju spora itd. U povoljnim uslovima bakterije se povećavaju i spremaju se za deobu. Lag faza može trajati od 10 minuta do nekoliko sati, ali u prosjeku iznosi 4-5 sati.
3. Faza logaritamskog ili eksponencijalnog rasta je period najintenzivnije podjele bakterija. Bakterije se dijele svakih 20-40 minuta. U ovoj fazi bakterije su posebno ranjive, što se objašnjava visokom osjetljivošću ćelija koje rastu na faktore okoline. Trajanje eksponencijalnog rasta ovisi o koncentraciji hranjivih tvari u supstratu i u prosjeku iznosi 5-6 sati.
5. Stacionarna faza rasta je uzrokovana postepenim iscrpljivanjem medijuma, akumulacijom litičkih enzima u njemu i hemijskom inhibicijom rasta mikrobnih ćelija metaboličkim produktima. Ova faza se razlikuje od prethodne po povećanju otpornosti bakterija na mnoge hemijske i fizičke faktore. Do početka ove faze, broj živih ćelija dostiže maksimalan nivo i ostaje na ovom maksimumu nekoliko sati, u zavisnosti od vrste mikroorganizama i karakteristika njihovog uzgoja. Na kraju ove faze, neki mikroorganizmi doživljavaju proces sporulacije.
6. Završnu fazu procesa reprodukcije – fazu starenja i umiranja – karakterizira umiranje bakterija uslijed iscrpljivanja hranljivog medija i nakupljanja metaboličkih produkata u njemu. Autoliza mikroorganizama se posmatra kao ekstremna manifestacija nestabilnosti ćelije nakon prestanka rasta. Trajanje ove faze može biti od nekoliko sati do nekoliko sedmica.
Datum objave: 2015-11-01; Pročitano: 2315 | Povreda autorskih prava stranice
studopedia.org - Studopedia.Org - 2014-2018 (0,002 s)…
Bakterije se, kao i svi živi organizmi, razmnožavaju. To se najčešće događa jednostavnom poprečnom podjelom u različitim ravnima. U tom slučaju nastaju različite kombinacije ćelija: uparene spojeve, pojedinačne ćelije, klasteri, lanci, paketi itd.
Neki mikroorganizmi se razmnožavaju sporulacijom (aktinomicete i gljive) i pupanjem (kvasac); neki mikroorganizmi se razmnožavaju spolno, ali većina ih se razmnožava aseksualno (vegetativno).
Pod povoljnim uslovima, reprodukcija se odvija izuzetnom brzinom - svakih 20-30 minuta matična bakterijska ćelija se deli na dve ćelije kćeri. Ćerka ćelija na kraju postaje matična ćelija i takođe se deli.
Dakle, podjela bakterija se odvija geometrijskom progresijom. Ako bi se takva podjela odvijala nesmetano, onda bi nakon 48 sati jedna bakterija mogla dati nastanak stotina milijardi ćelija, a nakon pet dana takvu masu koja bi ispunila bazene svih mora i okeana. Međutim, to se ne događa, jer na mikroorganizme utječu različiti faktori okoline.
Ćelijskoj diobi prethodi ravnomjerno povećanje ukupnog dušika, RNK i proteina u citoplazmi.
Tada dolazi do replikacije DNK (udvostručavanja). U ćeliji koja se dijeli, vodonične veze se prekidaju između spirala DNK i formiraju se jednostruke ćerke DNK spirale (slika 25).
25. Proces binarne fisije štapićastih prokariota
1 - formiranje pojedinačnih spirala DNK;
2 - udvostručavanje (replikacija) DNK;
3 - istezanje ćelije;
- formiranje septuma;
4 - završetak formiranja septuma i formiranje konveksnog ćelijskog zida;
5 - razdvajanje ćelija.
Neposredno nakon replikacije DNK, počinje produžavanje ćelije i formiranje poprečnog septuma zbog dva sloja citoplazmatske membrane koji strše jedan prema drugom.
Najčešće se u sredini matične ćelije formira septum, zbog čega ćelije kćeri imaju približno istu veličinu. Između slojeva septuma formira se ćelijski zid.
Jedinstvena spirala DNK u novim ćelijama služi kao šablon za stvaranje druge spirale, što rezultira formiranjem dvostruke spirale DNK sa obnovljenim vodikovim vezama i formiranjem novog nukleoida.
Tokom procesa reprodukcije, jedna od polovica ćelije stalno zadržava flagele.
U završnoj fazi razmnožavanja bakterija, flagele rastu u drugoj polovici.
Rast i razmnožavanje mikroorganizama zavisi od različitih faktora životne sredine i karakteristika vrste. Posmatranje razvoja mikroorganizama uzgajanih u tečnom hranljivom mediju u zatvorenim rezervoarima pokazuje da je za rast biomase potrebno prisustvo izvora energije, prisustvo komponenti neophodnih za sintezu biomase, odsustvo inhibitora u medijumu koji potiskuju rast ćelija i održavanje u okruženju potrebnih fizičkih i hemijskih uslova.
Pod ovim uslovima, rast mikroorganizama se može podeliti u nekoliko uzastopnih faza ili perioda (slika 26):
Rice. 26. Tipična krivulja rasta mikrobne populacije 1 - lag faza;
2 - faza ubrzanog rasta; 3 — faza logaritamskog (eksponencijalnog) rasta;
4 - faza usporavanja rasta; 5 - stacionarna faza rasta; 6 - faza starenja i smrti.
lag faza (eng. lag - kašnjenje) - period između sjetve bakterija i početka razmnožavanja. Tokom ovog perioda, bakterijska kultura se prilagođava hranljivom mediju. Manifestuje se u akumulaciji optimalne količine potrebnih enzima, u inaktivaciji nekog inhibitora prisutnog u medijumu, u klijanju spora itd. U povoljnim uslovima bakterije se povećavaju i spremaju se za deobu.
Lag faza može trajati od 10 minuta do nekoliko sati, ali u prosjeku iznosi 4-5 sati.
2. Faza ubrzanog rasta uočava se nakon lag faze i karakteriše je povećanje brzine diobe mikroorganizama i akumulacije biomase.
3. Faza logaritamskog ili eksponencijalnog rasta je period najintenzivnije podjele bakterija.
Bakterije se dijele svakih 20-40 minuta. U ovoj fazi bakterije su posebno ranjive, što se objašnjava visokom osjetljivošću ćelija koje rastu na faktore okoline. Trajanje eksponencijalnog rasta ovisi o koncentraciji hranjivih tvari u supstratu i u prosjeku iznosi 5-6 sati.
4. Faza usporavanja rasta je prijelazni period od eksponencijalnog rasta u stacionarnu fazu rasta. U ovoj fazi, hranjive tvari supstrata se iscrpljuju i u njemu se nakupljaju metabolički produkti, što smanjuje intenzitet razmnožavanja mikroorganizama.
Stacionarna faza rasta uzrokovana je postepenim iscrpljivanjem medijuma, akumulacijom litičkih enzima u njemu i hemijskom inhibicijom rasta mikrobnih ćelija metaboličkim produktima. Ova faza se razlikuje od prethodne po povećanju otpornosti bakterija na mnoge hemijske i fizičke faktore. Do početka ove faze, broj živih ćelija dostiže maksimalan nivo i ostaje na ovom maksimumu nekoliko sati, u zavisnosti od vrste mikroorganizama i karakteristika njihovog uzgoja.
Na kraju ove faze, neki mikroorganizmi doživljavaju proces sporulacije.
6. Završnu fazu procesa reprodukcije – fazu starenja i umiranja – karakterizira umiranje bakterija uslijed iscrpljivanja hranljivog medija i nakupljanja metaboličkih produkata u njemu. Autoliza mikroorganizama se posmatra kao ekstremna manifestacija nestabilnosti ćelije nakon prestanka rasta.
Trajanje ove faze može biti od nekoliko sati do nekoliko sedmica.
Datum objave: 2015-11-01; Pročitano: 2316 | Povreda autorskih prava stranice
studopedia.org - Studopedia.Org - 2014-2018 (0.001 s)…
Reprodukcija mikroorganizama je binarna podjela jednoćelijskih mikroorganizama (bakterije, rikecije, protozoe, kvasac), kao rezultat čega se formiraju dvije nove kćeri punopravne jedinke, obdarene genetskom informacijom matične stanice. Gljive slične kvascu mogu se razmnožavati pupanjem i sporama; Plijesni i aktinomiceti se obično razmnožavaju sporama.
Bakterije
Razmnožavaju se jednostavnom poprečnom podjelom.
Bakterije su haploidne ćelije. Sastav bakterijske ćelije uključuje kapsulu, ćelijski zid, citoplazmatsku membranu, citoplazmu u kojoj se nalaze mezozomi, ribosomi, nukleoid i inkluzije. Neke bakterijske ćelije imaju flagele i formiraju spore.
Za razliku od životinjskih ćelija, unutrašnje strukture bakterijske ćelije kao što su mezozomi, ribosomi i nukleoidi nemaju membrane koje ih odvajaju od citoplazme.
Prema načinu ishrane bakterije se dijele na autotrofe i heterotrofe, a prema načinu disanja - na aerobe i anaerobe.
Actinomycetes
Razmnožavaju se sporama i poprečnom podjelom (vezanjem) hifa.
Zauzimaju srednju poziciju između gljivica i bakterija. Među blistavim gljivama postoji mreža patogenih vrsta - uzročnika aktinomikoze. Mnogi aktinomiceti su proizvođači antibiotika. (cm.
antibiotici). U Bergeyjevom vodiču aktinomicete se nazivaju streptomiceti.
Kvasac
Postoje 2 vrste razmnožavanja kvasca - vegetativno (aseksualno) i spolno sa stvaranjem spora. Kod većine vrsta kvasca vegetativna reprodukcija se odvija pupanjem, rijetko diobom (Schizosaccharomyces). Asporogen. Kvasac se razmnožava samo pupanjem. Do polnog razmnožavanja dolazi u nepovoljnim uslovima kada kvasac prestane da pupi i pređe u vrećice (asci) sa sporama – askosporama.
Seksualni proces sastoji se od kopulacije (fuzije) 2 vegetativne ćelije tako što se spajaju i formiraju kopulacioni kanal, u kojem dolazi do fuzije delova plazme i jezgra ćelije, nazvane kariogamija, uz formiranje diploidnog zigota. , predstavlja 2 ćelije povezane kopulacionim kanalom.
Redukciona dioba, odnosno mejoza, praćena prepolovljenjem broja hromozoma, nastaje odmah, bez seksualnog procesa, a zigota se pretvara u ascus sa 4 haploidne spore, pa je vegetativna generacija takvih spora haploidna. Spore klijaju bez kopulacije. Ovako dolazi do razmnožavanja kvasca Zygosaccharomyces. Kod kvasca Saccharomyces, polni proces nastaje kada se spore ili ćelije koje su iz njih proklijale spoje i formiraju diploidnu zigotu, koja odmah počinje da pupi, formirajući diploidno potomstvo.
Mejoza se javlja neposredno prije formiranja spora.
Kalupi
Gljive razlikuju vegetativno, spolno i aseksualno razmnožavanje.
Vegetativno razmnožavanje može se vršiti odvajanjem dijelova micelija iz glavne mase, koji se mogu samostalno razvijati, kao i pupanjem micelija ili pojedinačnih ćelija gljiva kvasca.
Seksualna reprodukcija se sastoji od fuzije zametnih stanica, što rezultira stvaranjem zigote.
Aseksualna reprodukcija se provodi pomoću posebnih formacija koje se nazivaju spore. Spore se mogu razviti unutar posebnih posuda za spore ili na krajevima posebnih micelijskih izraslina - konidiofora.
Glavni način na koji se plijesni razmnožavaju je putem spora. Plijesan raste neverovatno brzo.
U običnoj plijesni kruha možete razlikovati male crne točkice - sporangije, u kojima se formiraju spore. Jedan sporangijum sadrži do 50.000 spora, od kojih je svaka sposobna da reprodukuje stotine miliona novih spora u samo nekoliko dana! A ako su uslovi povoljni, plijesan će se brzo pojaviti na knjizi, cipelama ili na srušenom drvetu u šumi.
Bakterije: Životnu aktivnost bakterija karakterizira rast- formiranje strukturnih i funkcionalnih komponenti ćelije i povećanje same bakterijske ćelije, kao i reprodukcija- samoreprodukcija, što dovodi do povećanja broja bakterijskih ćelija u populaciji.
Bakterije se razmnožavaju binarnom fisijom na pola, rjeđe pupanjem.
Aktinomicete, poput gljiva, mogu se razmnožavati sporama. Za jednu grupu jednoćelijskih cijanobakterija opisana je višestruka fisija (serija brzih uzastopnih binarnih fisija koje dovode do formiranja 4 do 1024 nove ćelije). Aktinomicete, kao razgranate bakterije, razmnožavaju se fragmentacijom filamentoznih ćelija.Gram-pozitivne bakterije se dijele urastanjem sintetiziranih diobenih septa u ćeliju, sintetiziraju poprečni septum od periferije prema centru uz učešće mezozoma.
i gram-negativne - sužavanjem (na mjestu diobe detektira se postepeno rastuća zakrivljenost CPM-a i stanične stijenke prema unutra), kao rezultat formiranja figura u obliku bučice, od kojih se formiraju dvije identične ćelije. Kada pupaju, pupoljak se formira i raste na jednom od polova matične ćelije; matična ćelija pokazuje znakove starenja i obično ne može proizvesti više od 4 ćelije kćeri.
Kod drugih bakterija, osim reprodukcije, opaža se i seksualni proces, ali u najprimitivnijem obliku.
Seksualni proces bakterija razlikuje se od seksualnog procesa eukariota po tome što bakterije ne stvaraju gamete i ne dolazi do fuzije stanica. Međutim, najvažniji događaj seksualnog procesa, odnosno razmjena genetskog materijala, također se događa u ovom slučaju. To se zove genetska rekombinacija.
Ćelijskoj diobi prethodi replikacija bakterijskog hromozoma prema polukonzervativnom tipu (dvolančani lanac DNK se otvara i svaki lanac dovršava komplementarni lanac), što dovodi do udvostručavanja molekula DNK jezgra bakterije - nukleoida. . Replikacija DNK odvija se u tri faze: inicijacija, elongacija ili rast lanca i završetak.
Reprodukcija spiroheta: poprečna fisija - podjela ćelija u bakterijama u kojoj matična stanica daje dvije kćeri ćelije. Izvodi se u tri faze:
1) replikacija molekula DNK kružnog hromozoma vezanog za mezozom, koji je takođe podeljen na dva dela;
2) oplemenjivanje dva ćerka prstenasta hromozoma korišćenjem mezozoma;
3) razdvajanje citoplazme poprečnom pregradom, koja se formira od periferije do centra ćelije.
Razmnožavanje gljiva:
Većina gljiva je sposobna za vegetativno, aseksualno i spolno razmnožavanje.
Karakterističan je pleomorfizam - prisutnost nekoliko vrsta sporulacije u isto vrijeme, na primjer, aseksualne i seksualne.
Vegetativno razmnožavanje
- Dijelovi micelija.
- Specijalizovane formacije: artrospore (oidije) sa tankim zidovima ili klamidiospore sa debelim zidovima, nastaju, uz određene razlike, kada se micelij raspada na delove, a zatim nastaje novi.
- Pupanje hifa ili pojedinačnih ćelija (na primjer, kod kvasca).
Takođe pupaju askospore kod torbara i bazidiospore u čamcima. Nastali pupoljci se postupno odvajaju, rastu i na kraju sami počinju da pupaju.
Aseksualna reprodukcija
Sama aseksualna reprodukcija se odvija putem spora.
U zavisnosti od načina formiranja, razlikuju se endogene i egzogene spore.
- Endogeni sporovi(sporangiospore) su karakteristične za niže gljive.
Formiraju se unutar posebnih ćelija zvanih sporangije.
- Egzogene spore obično se nazivaju konidiji, nalaze se u višim i nekim nižim gljivama.
Formiraju se na vrhovima ili stranama posebnih hifa - konidiofora, okomito orijentiranih, koji mogu biti jednostavni ili razgranati.
Prekriveni su gustom školjkom, pa su prilično stabilni, ali nepomični. Mogu se pokupiti zračnim strujama ili životinjama i prenijeti na značajne udaljenosti. Tokom klijanja nastaju rastna cijev, a zatim i hife.
Seksualna reprodukcija

Konjugacija gameta
Niže gljive karakteriziraju fuziju haploidnih gameta kroz izogamiju, anizogamiju (heterogamiju) ili oogamiju.
U slučaju oogamije razvijaju se genitalije - oogonia(žensko) i antheridia(muško). Tokom oplodnje dolazi do formiranja oospore- ovo je zigot koji je prekriven debelom ljuskom, provodi neko vrijeme u stanju mirovanja, nakon čega klija.
Brzina i faze razmnožavanja bakterija u stacionarnim uslovima.
Kada se bakterije uzgajaju na tekućem hranjivom mediju, uočava se donji, difuzni ili površinski (u obliku filma) rast kulture.
Rast šaržne kulture bakterija uzgojenih u tekućem hranjivom mediju podijeljen je u nekoliko faza, odnosno perioda:
1. faza kašnjenja;
2. logaritamska faza rasta;
3. faza stacionarnog rasta, odnosno maksimalna koncentracija bakterija;
4. faza bakterijske smrti.
Ove faze se mogu grafički prikazati u obliku segmenata krivulje bakterijske reprodukcije, odražavajući ovisnost logaritma broja živih stanica o vremenu njihovog uzgoja.
Lag faza je period između sjetve bakterija i početka razmnožavanja.
Trajanje lag faze je u prosjeku 4-5 sati.U isto vrijeme, bakterije se povećavaju u veličini i pripremaju se za podjelu; povećava se količina nukleinskih kiselina, proteina i drugih komponenti.
Logaritamska (eksponencijalna) faza rasta je period intenzivne diobe bakterija. Njegovo trajanje je oko 5-6 sati.U optimalnim uslovima rasta, bakterije se mogu podijeliti svakih 20-40 minuta.
U ovoj fazi bakterije su najranjivije, što se objašnjava visokom osjetljivošću metaboličkih komponenti ćelije koja intenzivno raste na inhibitore sinteze proteina, nukleinske kiseline itd.
Zatim dolazi stacionarna faza rasta, u kojoj broj živih ćelija ostaje nepromijenjen, čineći maksimalan nivo (M-koncentracija). Njegovo trajanje se izražava u satima i varira ovisno o vrsti bakterija, njihovim karakteristikama i uzgoju.
Proces rasta bakterija završava fazom smrti, koju karakterizira smrt bakterija u uvjetima iscrpljivanja izvora hranjivog medija i akumulacije u njemu metaboličkih produkata bakterija. Njegovo trajanje se kreće od 10 sati do nekoliko sedmica. Intenzitet rasta i razmnožavanja bakterija zavisi od mnogih faktora, uključujući optimalan sastav hranljive podloge, redoks potencijal, pH, temperaturu itd.
Brzina rasta bakterija ovisi i o vanjskim uvjetima i o fiziološkim karakteristikama same ćelije.
Pod povoljnim uslovima, rast bakterijske ćelije završava se razmnožavanjem. Glavni način na koji se većina bakterija razmnožava je jednostavno dijeljenje ćelije na pola. Podjeli prethodi replikacija (udvostručavanje) hromozoma. Ova dva procesa su usko povezana. Učestalost replikacije je regulirana brzinom rasta stanica. Replikacija bakterijskog hromozoma se vrši na prethodno opisan način (videti odeljak 3.2.5).
Proučavanje obrasca uniformne distribucije genetskog materijala između ćelija kćeri nastalih kao rezultat deobe matične ćelije omogućilo je G. Jacobu, S. Brenneru i T. Cousin (1963) da formulišu koncept replikona. Replikon je jedinica replikacije; to je dio DNK koji sadrži regulatorne elemente neophodne za nezavisnu replikaciju. Kod bakterija su to hromozomi i plazmidi. Svaki replikon sadrži najmanje dva lokusa uključena u kontrolu replikacije: strukturni replikatorski gen (inicijatorski gen), koji određuje sintezu proteina inicijatora, i posebno mjesto replikatora, koje prepoznaje signale za početak udvostručavanja hromozoma.
Nakon određenog perioda rasta, ćelija dostiže određeno fiziološko stanje. Od citoplazmatske membrane replikon prima signale o potrebi replikacije hromozoma i spremnosti ćelije za podjelu. Pod uticajem signala aktivira se aktivnost strukturnog gena i sintetiše se protein inicijator.
Djeluje na replikator i započinje replikaciju.
Postoji koordinirana interakcija između sistema replikacije hromozoma i deobe ćelije: deobi ćelije uvek prethodi duplikacija hromozoma. Kada se replikacija završi, počinje proces diobe ćelije. Kod gram-pozitivnih bakterija i cijanobakterija to se postiže formiranjem poprečnog septuma koji dijeli matičnu ćeliju na dvije jednake ćelije kćeri.
Podjela se odvija na sljedeći način.
Na početku
sintetizira se dvoslojna citoplazmatska membrana. Tada se na unutrašnjoj strani ćelijskog zida formiraju dva tuberkula. Oni intenzivno rastu i, prstenasto prodirući unutar ćelije između slojeva formirane citoplazmatske membrane, formiraju dvostruku pregradu koja dijeli ćeliju na pola.
Podjela gramatički najpažljivijih bakterija
nastaje stezanjem. U ovom slučaju, genomi divergiraju duž polova ćelije, citoplazmatska membrana i ćelijski zid se protežu, invaginirajući od periferije do centra ćelije dok ne dođu u kontakt jedan s drugim. Kao rezultat, ćelija je spojena u dvije kćeri ćelije. Podjela stanica formiranjem septuma ili konstrikcije naziva se binarnom zbog formiranja dvije identične ćelije kćeri.
Pored opisane binarne fisije, bakterije imaju još jednu poznatu metodu razmnožavanja: pupanje. Bakterije iz rodova Hyphomicrobium, Pedomicrobium i drugih, udružene u grupu pupajućih bakterija, razmnožavaju se pupanjem.
Ovi organizmi imaju izgled izduženih štapića (0,5x 2 µm), ponekad kruškolikog, koji završavaju hifama ili prostekama (izraslinama).
Reprodukcija kod ovih bakterija počinje formiranjem pupoljka na kraju hife ili direktno na matičnoj stanici.
Pupoljak izrasta u ćeliju ćerku, formira flagelum i odvaja se od matične ćelije. Po dolasku u zrelo stanje, flagelum se gubi i proces razvoja se ponavlja.
Za razliku od binarne fisije, tokom pupanja, originalna ćelija ostaje matična ćelija, a novonastala ćelija ostaje ćelija kćer.
Između njih postoje morfološke i fiziološke razlike.
Aktinomiceti se razmnožavaju fragmentima micelija i sporama. Kod nekih (rod Micromonospora) na hifama vegetativnog micelija nastaju pojedinačne spore, kod drugih (rod Streptomyces i dr.) na krajevima hifa zračnog micelija nastaju lanci spora, tzv. nazvani konidiofori.
Fragmenti micelija i spora klijaju u povoljnim uslovima vlažnosti i temperature i daju nove organizme.
Filamentozne cijanobakterije, pored binarne fisije, razmnožavaju se odsjecima trihoma i hormona. Potonji su skraćeni filamenti koji se sastoje od malih vegetativnih ćelija istog oblika i veličine. Kada srednje ćelije trihoma (niti) odumru, hormon izmiče iz ovojnice matičnog trihoma, raste, dijeli se, formirajući nove trihome.
Hormogonije, za razliku od majčinog trihoma, nemaju heterociste i nikada nisu okružene omotačem.
Bez obzira na to na koji način se odvija proces razmnožavanja bakterija, brzina ovog procesa je ogromna: za 24 sata može se promijeniti onoliko generacija koliko se čovjek ima za pet hiljada godina.
Brzina razmnožavanja ovisi o mnogim uvjetima i različita je za svaku vrstu bakterija. Ako su u mediju prisutne potrebne hranljive materije, temperatura i kiselost podloge su povoljne, podela svake ćelije se može ponoviti nakon 20-30 minuta (E. coli). Ovom brzinom reprodukcije, 472 * 1019 ćelija (273,72 generacije) može se formirati iz jedne ćelije dnevno.
Intenzivna reprodukcija je od velike biološke važnosti za bakterije. Osigurava očuvanje mikroorganizama na površini zemlje. Kada se pojave nepovoljni uslovi, oni masovno umiru, ali dovoljno je da negdje preživi nekoliko ćelija, a pod pogodnim uslovima iz njih će nastati veliko potomstvo ćelija.
Veličina populacije mikroorganizama u prirodnim staništima, na primjer, u tlu ili vodi, stalno se mijenja u skladu s promjenjivim životnim uvjetima.
Ali u laboratorijskim uslovima na hranljivim podlogama, promene u populaciji mikroorganizama se javljaju prirodno.
I također u odjeljku "REPRODUKCIJA BAKTERIJA"
Actinomycetes(Actinomyces) u prijevodu s latinskog je blistava gljiva, posebna grupa mikroorganizama koja ima niz morfoloških karakteristika nižeg tipa gljive i bakterije koja ne stvara spore.
Morfologija aktinomiceta
Struktura aktinomicita ima slične karakteristike sa filamentoznim gljivama, micelijski supovi imaju prosječnu debljinu od 0,7 mikrona, koja varira između 0,5-1,2 mm, što je mnogo manje nego kod gljiva.
Ravne ili blago zakrivljene niti koje nemaju poprečne pregrade karakteriziraju monopodijalno ili, u nekim slučajevima, navojno grananje. Sastav ćelijske membrane ima niz karakteristika gram-pozitivnih bakterija.
Reprodukcija aktinomiceta
Actinomyces se razmnožava pomoću supstratnog micelija koji klija u supstratu i zračnog micelija koji raste iz plodonosnih sporangiofora.
Voćke, u zavisnosti od vrste, imaju različite oblike kovrdža, od uvijenih do ravnih ili valovitih.
Neke vrste aktinomiceta imaju grane koje nose spore smještene u obliku kolutova ili snopova; često vise monopodijalno na nitima micelija.
Formiranje spora nastaje fragmentacijom ili segmentacijom.
Fragmentacija- ovo je proces drobljenja protoplasta grane koja nosi spore u stotinu ili više malih grudica koje sadrže bazofilne i nuklearne tvari.
Grudice, koje se pretvaraju u spore, nalaze se u dugom lancu u nosaču spora.
Segmentacija- ovo je proces podjele nosioca spora na segmente u obliku štapa, uz pomoć poprečnih pregrada, oni se zaokružuju i pretvaraju u spore.
Ljuske spora različitih vrsta imaju glatku, ponekad kvrgavu, nazubljenu, bodljikavu, dlakavu površinu. Izrasline na površini školjki jasno su vidljive kroz elektronski mikroskop.
U većini slučajeva aktinomiceti su aerofili i mezofili, ali su i termofili postali široko rasprostranjeni; mnoge njihove vrste su sposobne proizvoditi pigmente različitih boja.
Aktinomiceti, koji imaju raznolik skup enzima, u stanju su sintetizirati različite tvari i otpuštati ih u velikim količinama u okoliš. Među ovim supstancama sa visokom fiziološkom aktivnošću su mnogi vitamini, neke aminokiseline, toksini, karotenoidi, fitohormoni i drugi.
Vrijedi spomenuti i sposobnost aktinomiceta da formiraju različite vrste antibiotika.







